Highlighting the article by in the May 2015 issue of Insectes Sociaux
Written by Aniruddha Mitra
Mate recognition is a vital aspect of communication, essential for survival of the species, as in nature males and females have to find each other and recognize the sex of the partner before engaging in reproduction related behaviors and mating. Sensing chemicals form a major part of the sensory world of insects, and chemical signals (pheromones) released by an individual are perceived by others, following which, the kind of signal received alters the behavior and decision making of the receiver.
Chemical signals called sex pheromones play an important role in attracting mates, sex recognition and mating related behaviors in the majority of insects. In many insects, sex pheromones are volatile chemicals that serve to attract mates from a distance. In some insects non-volatile chemicals like hydrocarbons present on the body surface have been implicated as sex pheromones. The external body surface (cuticle) of insects is covered by a waxy layer of hydrocarbons, which primarily serves to provide protection from dessication and pathogens. However in some insects, especially the social Hymenoptera (ants, social wasps and social bees), the cuticular hydrocarbons (CHCs) also play a role in communication.
CHCs can provide several types of information about an individual ant, bee or wasp. Variations in the composition of CHCs, allow workers to identify their queen, distinguish members of their own nest from those of other nests as well as obtain other forms of information about an individual. CHCs may also function as sex pheromones, and differences in CHC profiles of males and females (sexual dimorphism in CHCs) have been discovered in diverse groups of insects (more than 100 species from 7 different orders).
The social hymenopteran insects have been the focus of numerous studies of CHCs and their role in communication. However, few studies have looked at sexual dimorphism of CHCs or role of CHCs in mate recognition in social hymenopterans. Also sex pheromones (including CHCs as well as other volatile chemicals) remain a relatively less known area in the biology of Hymenoptera. Hence to generate more knowledge in this area, we investigated the question of whether sexual dimorphism of CHCs exists in our study species, a social wasp from southern India. We also tried to detect potential volatile cues that may attract mates from a distance.
The wasp Ropalidia marginata has been studied over more than two decades, and only in the last 5 years have we uncovered aspects of chemical communication in this species. We have discovered that the queen signals her presence to workers through a gland secretion and that through CHCs, workers can differentiate their own nest members from members of other nests. In our work detailed in Insectes Sociaux, we investigated another aspect of possible chemical communication in this species – mate recognition through chemical cues.

Fig. 1: Male (left) and female (right) R. marginata wasps.
R. marginata wasps live in colonies comprising of a queen and several workers and build nests of paper from cellulose from plants. There is no seasonal variation in colony cycle, and all females can mate and have the potential to become a queen. Males stay on the nest for about a week and then leave the colony to live a solitary nomadic life. Mating is believed to occur on trees and other areas where females go out to collect food and building material, and has never been seen on the nest. Before mating, males approach females and touch them with their antennae, following which the male mounts the female and grasps her antennae with his antennae, finally culminating in mating.
Since the male and female touch each other with their antennae before mating, we looked for any non-volatile cue that may be present on the cuticle and be perceived by touching with antennae. We analyzed the chemical composition of non-volatile CHCs by gas chromatography (see Fig. 2). For a CHC to act as sex pheromone, there should be some consistent difference between the CHC profiles of males and females, otherwise the wasps would not be able to differentiate between the two. We did not find any significant differences in CHC composition between males and females, therefore we ruled out the involvement of any non-volatile CHC as a mate recognition cue.

FIg. 2: A representation of the gas chromatography (GC) device that detects the chemical components present in the CHCs of male and female wasps. We found no significant differences in the CHC chemical profiles between male and female wasps.
Next, we explored the possibility that there are volatile cues that may act as sex attractants. We then further analysed the CHCs in order to detect chemicals that may be volatile. This analysis showed that volatile chemicals are absent in the CHCs. Finally we did a behavioral assay to see if males and females are attracted towards each other from a distance. By doing this, we could test for any volatile compound from the cuticle that may have been missed in the chemical analysis, and would also cover volatile cues from sources other than CHCs. Wasps were introduced at the center of a T-tube maze (see Fig. 3), and a female and a male were kept at either end of the T-tube (separated from the test wasp by a mesh). We found that males did not spend more time towards the side of the T-tube containing a female, compared to the side that had a male. The same held true for females, who did not spend more time towards the male side, compared to the female side. Thus we failed to find evidence of any volatile cues that may attract members of the opposite sex towards each other.
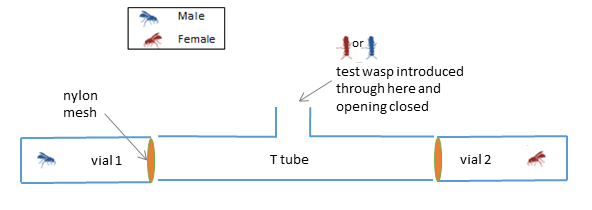
Fig. 3: A representation of the T-tube choice apparatus used to test whether female and male wasps are attracted to a member of the opposite sex that they can smell, but cannot see. Neither female nor male wasps showed a preference for either the same or the opposite sex.
It remains unclear whether any sex pheromone exists in this species. In Hymenoptera, apart from CHCs, sex pheromones have been reported from various glands, so it is possible that some glandular secretion that is released after the male and female come in contact with each other plays a role in mate recognition. Compounds other than hydrocarbons like peptides and proteins are also reported to be present on the cuticle, and these may have a role in mate attraction. Finally it may also be possible that mate recognition in this species does not involve chemical cues and involves visual cues instead. Some social wasps have the ability to recognize faces (see, for example, here) and since the face of males is yellow, while those of females is brown (see Fig. 1), it is possible that they might use this visual cue to recognize members of the opposite sex, but we still need to search for the answer to how male and female R. marginata manage to find and recognize each other in a large and complex environments.
About the author: Aniruddha Mitra is a researcher at Laboratoire Evolution, Génomes, Comportement, Ecologie, CNRS, Gif sur Yvette, France. He can be reached by email at mitra.aniruddha@gmail.com.



![[Dorylus Male] Male ants also fly, and some, like this Dorylus driver ant from a Ugandan rainforest (shown with the author), are among the largest ants in the world (photo by Alex Wild).](https://insectessociaux.com/wp-content/uploads/2015/05/dorylus-male.jpg)
